Zika Virus (ZIKV) was first isolated in 1947 from a sentinel monkey in Uganda and despite sporadic local outbreaks only caused mild disease in humans. The emergence of ZIKV combined with severe pathogenicity in South America as early as 2019 however raised questions about the molecular evolution of ZIKV since ZIKV was previously only associated with arthralgia and a mild febrile illness but not neuropathological disorders including abnormal foetal brain development and Guillan-Barre Syndrome (GBS).
ZIKV is a flavivirus closely related to Dengue Virus (DENV) with single stranded positive stranded RNA genome of approximately 10800 bp. Similar to DENV, the ZIKV RNA encodes for a single polyprotein that it is cleaved into the structural (Capsid (C), pre-membrane (prM), and envelope (E)) and non-structural (NS1, NS2A, NS2B, NS3, NS4A, 2K, NS4B, and NS5) proteins with the replication taking place in the cytoplasm of infected although at least the C and NS5 proteins localise to the nucleolus and to nuclear speckles respectively, suggesting that the nuclear localisation of these proteins might be required for efficient viral replication.
Based on genetic and phylogenetic analyses, two lineages can be identified, with epidemics in Micronesia/Yap in 2007, French Polynesia 2019-2019 and the current epidemic in South America being linked to the Asian lineage. Phylogenetic analyses of all contemporary strains share greater homology to ZIKV P6-740 strain isolated from mosquitoes in Malaysia in 1966 rather than the ZIKV IbH-30656 isolated from Nigeria in 1968, suggesting that the current strains circulating in the Pacific and South America are rooted in the African lineage but are descendants of the ZIKV Malaysia/1966. Further analysis revealed that the current Brazilian strains are closer related to ZIKV H/PF/2019 (ZIKV French Polynesia/2019) rather than ZIKV FSM (Federal States of Micronesia)/2007. Since the ZIKV outbreak in Polynesia but not in Micronesia has been associated with an increased risk of GBS and of microcephaly, it might be possible that ZIKV H/PF/2019 and the descendants isolated in the Americas exhibit mutations that cause foetal brain abnormalities and increase the risk of developing GBS. In order to investigate if ZIKV H/PF/2019 is indeed more pathogenic than ZIKV FSM/2007 recently developed mouse models might provide be useful. In addition, the use of the recently developed infectious clone of ZIKV FSS1305 –a strain isolated in Cambodia in 2010- might aid these studies by introducing mutations into viral genes that can be found in isolates from the Americas and/or from French Polynesia. The use of historical strains including the original ZIKV MR-766 isolate to study ZIKV related pathogenesis however is problematic since these strains were originally propagated in mice brains and are therefore highly adapted whereas modern isolates are propagated in mosquitoe C6/36 and Vero cells, although historically the repeated passage of ZIKV MR-766 in mice brain yielded important insights in ZIKV pathogenesis. Using mice however as a model system for studying the teratogenicity of ZIKV is limited since offspring of immunocompetent mice does not develop microcephaly, suggesting that in humans other factors besides ZIKV infection are required for the development of microcephaly. One of the contributing factors might be a previous infection or vaccination against DENV, others might be co-infection with other pathogens that allow ZIKV or ZIKV infected immune cells to cross the placental barrier and infect the embryo.
In the past months, the infection of brain organoids and human neural precursor cells (hNPC) with ZIKV not only let to the identification of the receptor that allows ZIKV entry into neural cells (Axl) but also suggest how ZIKV induces abnormal brain development.
As discussed in detail in previous posts, the infection of hNPC and brain organoids downregulates the expression of genes regulating multiple cellular pathways such as DNA repair, DNA replication and cell cycle progression, suggesting that apoptosis is induced by stalled replication forks and mitotic defects. In general, a model was proposed in which ZIKV induces the depolarisation of mitochondria and inducing caspase-3 dependent apoptosis. Upregulation of autophagy related genes (ATG) on the other might benefit viral replication by increasing both lipophagy and the formation of viral replication centres whilst at the same time contributing to autophagy mediated cell death.
Close analysis of mouse brains derived from immunodeficient SJL mice pups but not wt C57BL/6 born to females infected with a ZIKV strain isolated from Brazil (ZIKV BR) shows that cortical neurons as well as neurons derived from the thalamus and hypothalamus exhibit nuclear debris suggesting that ZIKV BR does induce apoptosis as well as exhibiting ocular defects similar to human neonate patients and anatomical changes similar to those seen in microcephaly in neonates. In a similar way, injection of mouse embryos with ZIKV SZ01 (Asian lineage isolated from a Chinese patient) at E13.5 leads to cell cycle arrest as measured by BrdU incorporation, apoptosis of neuronal cells and abnormal differentiation of neural precursor cells suggesting that neuronal death is a common feature of ZIKV infected pups. Furthermore, both ZIKV BR and ZIKV SZ01 replicates efficiently in brain tissue of immunodeficient mice despite not being adapted to mouse cells. In a separate study, injection of ZIKV SZ01 into pregnant C57BL/6 in utero (thus bypassing the placental barrier) affects the cortical development of offspring mice as early as 96 hrs p.i. concomitant with a downregulation of the expression of genes related to cell proliferation and the negative regulation of apoptosis whereas the expression of genes encoding for proteins involved in antiviral signaling is increased, suggesting that ZIKV induces both apoptotic and antiviral signaling pathways in embryonic cells.
The in uteroinfection of (mouse) foetal brains with ZIKV SZ01 also decreases the expression of proteins that previously have been linked to the development of microcephaly in particular those that are involved in the separation of chromosomes during metaphase and anaphase, suggesting that ZIKV infected embryonic and/or foetal cells might exhibit incomplete cytokinesis which may lead to the formation of micronuclei and subsequent apoptosis. In a similar way, hNPC infected with ZIKV MR766 exhibit a decrease in the expression of the very same genes confirming that the downregulation of genes regulating mitotic progression might contribute to the development of microcephaly in human foetuses and neonates. Interestingly, the infection of hNPC with ZIKV MR766 also induces the downregulation of Aurora Kinase B, suggesting that the induction of the NoCut checkpoint that regulates the abscission in response to chromosome segregation defects both in yeast and animal cells might be inhibited thus leading to mitotic defects.
 |
| Table: ZIKV induced increase of gene expression in E13.5 and E17.5 mice embryos |
The in uteroinfection of (mouse) foetal brains with ZIKV SZ01 also decreases the expression of proteins that previously have been linked to the development of microcephaly in particular those that are involved in the separation of chromosomes during metaphase and anaphase, suggesting that ZIKV infected embryonic and/or foetal cells might exhibit incomplete cytokinesis which may lead to the formation of micronuclei and subsequent apoptosis. In a similar way, hNPC infected with ZIKV MR766 exhibit a decrease in the expression of the very same genes confirming that the downregulation of genes regulating mitotic progression might contribute to the development of microcephaly in human foetuses and neonates. Interestingly, the infection of hNPC with ZIKV MR766 also induces the downregulation of Aurora Kinase B, suggesting that the induction of the NoCut checkpoint that regulates the abscission in response to chromosome segregation defects both in yeast and animal cells might be inhibited thus leading to mitotic defects.
 |
| Table: Downregulation of gene expression; genes related to microcephaly also regulate mitosis |
Another possibility is however that the infection of ZIKV prolongs S phase by the activation of intra S phase checkpoints and subsequently arresting infected cells in G2 rather than M phase. This notion is supported by flow cytometry analysis of ZIKV MR766 infected hNPC as well as by measuring the presence of Histone H3-P (Ser10), a marker that allows the differentiation between G2 and M phase by flow cytometry in ZIKV SZ01 infected mouse embryonic cells. In addition to the observed G2 arrest, the latter also exhibited duplicated centrosomes suggesting that ZIKV infected indeed do not enter mitosis. As evidenced by the expression of Pax6, Sox2 and Tbr2, ZIKV mainly infects primary neuronal progenitor cells that undergo cell proliferation rather than differentiated neurons, although post mitotic neutrons may be infected as well unless ZIKV positive post mitotic neurons represent neurons that got initially infected prior differentiation and failed to undergo apoptosis prior differentiation. Alternatively, post mitotic neurons may be a infected as a result of viral spread following the infection of immature neurons.
In contrast to other previously published papers using isolates from Brazil of French Polynesia however, ZIKV SZ01 might cross the placental barrier since intraperitoneal injection of ZIKV SZ01 causes abnormal foetal brain development in C57BL/6 mice. Additionally, the results suggest that in C57BL/6 the injection of ZIKV i.p. might not cause viraemia as confirmed by previous studies but unfortunately, no side by side experiments with other ZIKV isolates were performed.
Since the infection of susceptible cells with ZIKV probably induces a TLR-3 dependent antiviral response and thus might induce bystander apoptosis, further research is needed to elucidate the precise signaling pathways that induce the activation of caspase-3. Here the use of specific inhibitors is warranted. In addition, the contribution of autophagy to cell death has not been established despite the reported upregulation of genes such as Bmf, Irgm1, Bcl-2, Htt, Abl1 and Casp6 that are involved in the regulation of the formation of autophagosomes. Since the overexpression of some of the proteins –especially Bcl-2 and Bmf- inhibits rather than promotes the formation of autophagosomes whereas the overexpression of Irgm-1 promotes neuronal autophagy and survival of mouse neurons, the connection between ZIKV induced autophagy and apoptosis might be more complex. Adding to the complexity, the upregulation of Casp6 however might prevent p62/SQSTM-1 dependent selective autophagy despite the upregulation of Htt by cleaving Htt. In addition, cleavage of Beclin-1 by caspase-3 -which generates a C-terminal fragment of Beclin-1- might also inhibit the formation of autophagosomes.
 |
| Figure: HTT recognises p62/SQSTM1 and is involved in the recruitment of the ULK complex |
Interestingly, A549 cells infected with ZIKV H/PF/2019 do not exhibit autophagosomes or autophagosome-like vesicles whereas human skin fibroblasts infected with ZIKV MR766 do. Both neurons and NPC infected with ZIKV BR at an MOI of 10 exhibit large viral protein aggregates at 24 hrs p.i. suggesting the presence viral replication centres whose precise nature is not known.
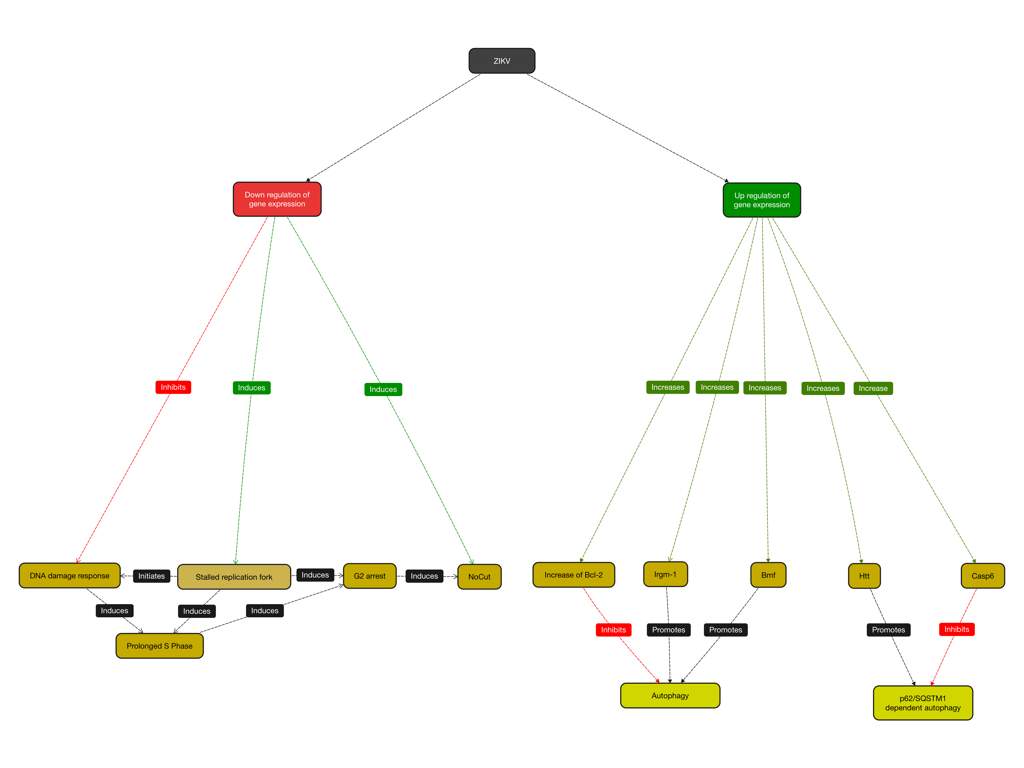 |
| Figure: Hypothetical network in which ZIKV induces the NoCut pathway and regulates autophagy |
ZIKV NS5 and STAT2: mitotic defect?
The NS5 protein derived from various members of the Flaviviridae including DENV, WNV, Spondweni Virus, YFV, JEV, Langat Virus (LGV), Yokose Virus (YOKV) and Usutu Virus (USUV) has been shown to bind and degrade STAT2 and thus abrogate the type I Interferon response in Vero cells. In a similar way, the expression of ZIKV NS5 prevents the nuclear translocation of STAT2 by promoting the degradation of STAT2 via ubiquitinylation and subsequent targeting to proteasomes both in transfected and ZIKV Fortaleza infected cells in a species dependent manner, e.g.in Vero, primary human fibroblasts and HEK 293T cells but not in primary mouse fibroblasts derived from IFNAR -/- mice that are otherwise permissible for ZIKV. Three features make NS5 an interesting protein. First, a subfraction of NS5 localises to nuclear speckles in both infected and NS5 transfected cells, suggesting that NS5 is imported into the nucleus where it might co-localise and interact with proteins such as the viral RNA dependent RNA polymerase (RdRp) at sites of active viral RNA synthesis. Second, overexpressed ZIKV NS5 co-localises with (overexpressed) STAT2 in a region that corresponds with the ER, suggesting that the ER stress might be involved in the degradation. Third, a subset of Vero cells expressing ZIKV NS5 exhibits aberrant mitosis reminiscent to those observed in Vero cells expressing the coronaviral N protein, which is characterised by the presence of a cleavage furrow. The activation of STAT2 has been shown to promote neuronal differentiation in human left cerebrum which includes the development of the visual perception. Since foetal ZIKV infection has been associated with visual defects, it might be possible that ZIKV mediated degradation of STAT2 not only abrogates type I IFN dependent signalling but also interferes with differentiation of neuronal cells, in particular those related to visual perception.
In conclusion, the past months saw tremendous progress in the development of mouse models allowing closer examination of the link of maternal ZIKV infection and the development of microcephaly due to abnormal development of the foetal brain in offspring. With the exception of one study, all of the models successfully employed involved the use of immunodeficient mice suggesting that the passage of ZIKV through the maternal placenta in immunocompetent animals is prevented in most circumstances. In addition to the animal model, studies using human neural progenitor cells and brain organoids suggest that in ZIKV infected cells the expression of genes controlling the cell cycle and inhibiting apoptosis is downregulated, data that are confirmed in transcriptome analysis of foetal brains of infected mice pups. In general however the decrease of genes expression is more pronounced in experimentally infected hNPC than in foetal brain which is probably due to antiviral signalling pathways that restrict the initial infection of embryonal cells in vivo.
Further reading
Wang L, Valderramos SG, Wu A, Ouyang S, Li C, Brasil P, Bonaldo M, Coates T, Nielsen-Saines K, Jiang T, Aliyari R, & Cheng G (2019). From Mosquitos to Humans: Genetic Evolution of Zika Virus. Cell host & microbe, 19 (5), 561-5 PMID: 27091703
Tang H, Hammack C, Ogden SC, Wen Z, Qian X, Li Y, Yao B, Shin J, Zhang F, Lee EM, Christian KM, Didier RA, Jin P, Song H, & Ming GL (2019). Zika Virus Infects Human Cortical Neural Progenitors and Attenuates Their Growth. Cell stem cell, 18 (5), 587-90 PMID: 26952870
Rodriguez-Morales AJ, Bandeira AC, & Franco-Paredes C (2019). The expanding spectrum of modes of transmission of Zika virus: a global concern. Annals of clinical microbiology and antimicrobials, 15 PMID: 26939897
Beckham JD, Pastula DM, Massey A, & Tyler KL (2019). Zika Virus as an Emerging Global Pathogen: Neurological Complications of Zika Virus. JAMA neurology PMID: 27183312
Miner JJ, Cao B, Govero J, Smith AM, Fernandez E, Cabrera OH, Garber C, Noll M, Klein RS, Noguchi KK, Mysorekar IU, & Diamond MS (2019). Zika Virus Infection during Pregnancy in Mice Causes Placental Damage and Fetal Demise. Cell, 165 (5), 1081-91 PMID: 27180225
Cugola, F., Fernandes, I., Russo, F., Freitas, B., Dias, J., Guimarães, K., Benazzato, C., Almeida, N., Pignatari, G., Romero, S., Polonio, C., Cunha, I., Freitas, C., Brandão, W., Rossato, C., Andrade, D., Faria, D., Garcez, A., Buchpigel, C., Braconi, C., Mendes, E., Sall, A., Zanotto, P., Peron, J., Muotri, A., & Beltrão-Braga, P. (2019). The Brazilian Zika virus strain causes birth defects in experimental models Nature DOI: 10.1038/nature18296
Deng YQ, Zhao H, Li XF, Zhang NN, Liu ZY, Jiang T, Gu DY, Shi L, He JA, Wang HJ, Sun ZZ, Ye Q, Xie DY, Cao WC, & Qin CF (2019). Isolation, identification and genomic characterization of the Asian lineage Zika virus imported to China. Science China. Life sciences, 59 (4), 428-30 PMID: 26993654
Driggers RW, Ho CY, Korhonen EM, Kuivanen S, Jääskeläinen AJ, Smura T, Rosenberg A, Hill DA, DeBiasi RL, Vezina G, Timofeev J, Rodriguez FJ, Levanov L, Razak J, Iyengar P, Hennenfent A, Kennedy R, Lanciotti R, du Plessis A, & Vapalahti O (2019). Zika Virus Infection with Prolonged Maternal Viremia and Fetal Brain Abnormalities. The New England journal of medicine PMID: 27028667
Li C, Xu D, Ye Q, Hong S, Jiang Y, Liu X, Zhang N, Shi L, Qin CF, & Xu Z (2019). Zika Virus Disrupts Neural Progenitor Development and Leads to Microcephaly in Mice. Cell stem cell PMID: 27179424 Wu KY, Zuo GL, Li XF, Ye Q, Deng YQ, Huang XY, Cao WC, Qin CF, & Luo ZG (2019). Vertical transmission of Zika virus targeting the radial glial cells affects cortex development of offspring mice. Cell research PMID: 27174054
Grant, A., Ponia, S., Tripathi, S., Balasubramaniam, V., Miorin, L., Sourisseau, M., Schwarz, M., Sánchez-Seco, M., Evans, M., Best, S., & García-Sastre, A. (2019). Zika Virus Targets Human STAT2 to Inhibit Type I Interferon Signaling Cell Host & Microbe DOI: 10.1016/j.chom.2019.05.009
Shan C, Xie X, Muruato AE, Rossi SL, Roundy CM, Azar SR, Yang Y, Tesh RB, Bourne N, Barrett AD, Vasilakis N, Weaver SC, & Shi PY (2019). An Infectious cDNA Clone of Zika Virus to Study Viral Virulence, Mosquito Transmission, and Antiviral Inhibitors. Cell host & microbe PMID: 27198478 Ashour J, Laurent-Rolle M, Shi PY, & García-Sastre A (2009). NS5 of dengue virus mediates STAT2 binding and degradation. Journal of virology, 83 (11), 5408-18 PMID: 19279106
Gelman A, Rawet-Slobodkin M, & Elazar Z (2019). Huntingtin facilitates selective autophagy. Nature cell biology, 17 (3), 214-5 PMID: 25720962
Amaral N, Vendrell A, Funaya C, Idrissi FZ, Maier M, Kumar A, Neurohr G, Colomina N, Torres-Rosell J, Geli MI, & Mendoza M (2019). The Aurora-B-dependent NoCut checkpoint prevents damage of anaphase bridges after DNA replication stress. Nature cell biology, 18 (5), 516-26 PMID: 27111841
Schröder-Heurich B, Wieland B, Lavin MF, Schindler D, & Dörk T (2019). Protective role of RAD50 on chromatin bridges during abnormal cytokinesis. FASEB journal : official publication of the Federation of American Societies for Experimental Biology, 28 (3), 1331-41 PMID: 24344331
Wang L, Huang J, Jiang M, & Lin H (2012). Signal transducer and activator of transcription 2 (STAT2) metabolism coupling postmitotic outgrowth to visual and sound perception network in human left cerebrum by biocomputation. Journal of molecular neuroscience : MN, 47 (3), 649-58 PMID: 22219046
Kumar A, Bühler S, Selisko B, Davidson A, Mulder K, Canard B, Miller S, & Bartenschlager R (2019). Nuclear localization of dengue virus nonstructural protein 5 does not strictly correlate with efficient viral RNA replication and inhibition of type I interferon signaling. Journal of virology, 87 (8), 4545-57 PMID: 23408610
Uchil, P., Kumar, A., & Satchidanandam, V. (2006). Nuclear Localization of Flavivirus RNA Synthesis in Infected Cells Journal of Virology, 80 (11), 5451-5464 DOI: 10.1128/JVI.01982-05
Davis, M., & Gack, M. (2019). Ubiquitination in the antiviral immune response Virology, 479-480, 52-65 DOI: 10.1016/j.virol.2019.02.033
No comments:
Post a Comment